



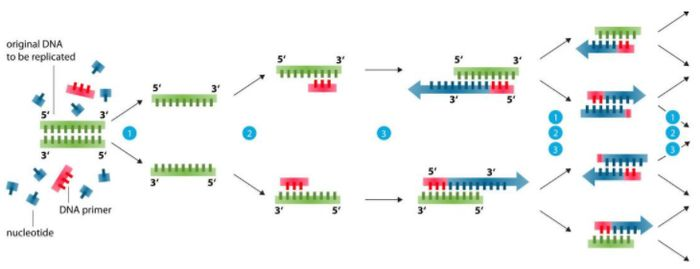
Základ návrhu základného náteru (99 % problémov je možné vyriešiť)
1. Dĺžka priméru: Učebnica vyžaduje 15-30bp, zvyčajne okolo 20bp.Skutočná podmienka je lepšia 18-24 bp, aby sa zabezpečila špecificita, ale čím dlhší, tým lepší, príliš dlhý primér tiež zníži špecificitu a zníži výťažok.
2. Rozpätie amplifikácie priméru: Vhodné je 200-500 bp a fragment sa môže za špecifických podmienok rozšíriť na 10 kb.
3. Základná báza: Obsah G+C by mal byť 40-60%, príliš malý amplifikačný efekt G+C nie je dobrý, pri príliš veľkom množstve G+C sa ľahko objavia nešpecifické pásy.ATGC sa najlepšie distribuuje náhodne, pričom sa vyhýba zhlukom viac ako 5 purínových alebo pyrimidínových nukleotidov.Multi-gc pre 5' koniec a medzisekvencie na zvýšenie stability, vyhnite sa bohatému GC na 3' konci, žiadnemu GC na posledných 3 bázach alebo žiadnemu GC na 3 z posledných 5 báz.
4. Vyhnite sa sekundárnej štruktúre v priméroch a vyhnite sa komplementácii medzi dvoma primérmi, najmä komplementácii na 3' konci, inak sa vytvorí primérový dimér a vytvoria sa nešpecifické amplifikované pásy.
5. Bázy na 3' konci primérov, najmä posledná a predposledná báza, by mali byť prísne spárované, aby sa predišlo zlyhaniu PCR v dôsledku nespárovaných koncových báz.
6. Priméry majú alebo môžu byť pridané s vhodnými miestami štiepenia a amplifikovaná cieľová sekvencia by mala mať prednostne vhodné miesta štiepenia, čo je veľmi prospešné pre analýzu štiepenia alebo molekulárne klonovanie.
7. Špecifickosť primérov: priméry by nemali mať žiadnu zjavnú homológiu s inými sekvenciami v databáze sekvencií nukleových kyselín.
8. Naučte sa používať softvér: PP5, Oligo6, DNAstar, Vector NTI, primer3 (Tento online dizajn funguje najlepšie).
Vyššie uvedený obsah môže vyriešiť najmenej 99 % problémov s návrhom základného náteru.
Ovládajte detaily návrhu základného náteru
1. Dĺžka základného náteru
Celková dĺžka základného náteru je 18~30 báz.Vo všeobecnosti je najdôležitejším faktorom určujúcim teplotu žíhania priméru dĺžka priméru.Teplota žíhania priméru je všeobecne zvolená (hodnota Tm -5 °C) a niektoré priamo používajú hodnotu Tm.Na približný výpočet teploty žíhania primérov možno použiť nasledujúce vzorce.
Keď je dĺžka priméru menšia ako 20 bp: [4(G+C)+2(A+T)]-5℃
Keď je dĺžka priméru väčšia ako 20 bp: 62,3 ℃ + 0,41 ℃ (% GC) -500/dĺžka - 5 ℃
Okrem toho sa na výpočet teploty žíhania dá použiť aj veľa softvérov, princíp výpočtu bude iný, takže niekedy môže mať vypočítaná hodnota malú medzeru.Na optimalizáciu PCR reakcií sa pre najlepšiu účinnosť a špecifickosť používajú najkratšie priméry, ktoré zaisťujú teplotu nasadenia najmenej 54 °C.
Celkovo sa špecificita priméru zvyšuje štvornásobne pre každý ďalší nukleotid, takže minimálna dĺžka priméru pre väčšinu aplikácií je 18 nukleotidov.Horná hranica dĺžky priméru nie je veľmi dôležitá, hlavne súvisí s účinnosťou reakcie.V dôsledku entropie platí, že čím dlhší je primér, tým nižšia je rýchlosť, pri ktorej dochádza k napojeniu na cieľovú DNA za vzniku stabilného dvojvláknového templátu pre väzbu DNA polymerázy.
Pri použití softvéru na navrhovanie primérov možno dĺžku primérov určiť postupne podľa hodnoty TM, najmä pre priméry fluorescenčnej kvantitatívnej PCR by sa mala kontrolovať TM = 60 °C.
2. Obsah GC
Vo všeobecnosti je obsah G+C v sekvenciách primérov 40 % až 60 % a obsah GC a hodnota Tm páru primérov by mali byť koordinované.Ak má primér závažnú tendenciu GC alebo AT, na 5' koniec priméru možno pridať príslušné množstvo A, T alebo G a C konca.
3. Teplota žíhania
Teplota žíhania by mala byť o 5 ℃ nižšia ako teplota uvoľnenia z reťaze.Ak je počet báz primérov malý, teplota anelácie sa môže primerane zvýšiť, čo môže zvýšiť špecificitu PCR.Ak je počet báz veľký, teplota žíhania sa môže primerane znížiť.Teplotný rozdiel nasadania medzi párom primérov 4 °C ~ 6 °C neovplyvní výťažok PCR, ale v ideálnom prípade je teplota nasadenia páru primérov rovnaká, ktorá sa môže meniť medzi 55 °C ~ 75 °C.
4. Vyhnite sa oblasti sekundárnej štruktúry amplifikačnej šablóny
Pri výbere amplifikovaného fragmentu je najlepšie vyhnúť sa oblasti sekundárnej štruktúry templátu.Stabilnú sekundárnu štruktúru cieľového fragmentu možno predpovedať a odhadnúť pomocou príslušného počítačového softvéru, ktorý je užitočný pri výbere šablóny.Experimentálne výsledky ukazujú, že expanzia je často neúspešná, keď je voľná energia (△G) oblasti, ktorá sa má expandovať, menšia ako 58,6 kJ/mol.
5. Nesúlad s cieľovou DNA
Keď je amplifikovaná cieľová DNA sekvencia veľká, primér sa môže viazať na viaceré časti cieľovej DNA, čo vedie k tomu, že sa vo výsledku objavia viaceré pásy.Tentokrát je potrebné použiť testovanie softvéru BLAST, webovú stránku:http://www.ncbi.nlm.nih.gov/BLAST/.Vyberte možnosť Zarovnať dve sekvencie (bl2seq).
Prilepenie sekvencií primérov do zóny 1 a cieľových sekvencií DNA do zóny 2 je vzájomne zameniteľné a BLAST vypočítava komplementárne, antisense a ďalšie možnosti, takže používatelia si nemusia všimnúť, či sú oba reťazce sense reťazce.Môžete zadať aj číslo GI, ak poznáte číslo GI sekvencie v databáze, takže nemusíte vkladať veľkú časť sekvencie.Nakoniec kliknite na Zarovnať na 3, aby ste zistili, či má primér viacero homológnych miest v cieľovej DNA.
6. Primer terminál
3 'koniec základného náteru je miesto, kde začína predlžovanie, takže je dôležité zabrániť tomu, aby sa tam začali nezhody.3' koniec by nemal byť viac ako 3 po sebe idúce G alebo C, pretože to spôsobí, že primér sa omylom spustí v oblasti sekvencie obohatenej G+C.3' koniec nemôže tvoriť žiadnu sekundárnu štruktúru, s výnimkou špeciálnych PCR (AS-PCR) reakcií, 3' koniec priméru nemôže byť nesprávne spárovaný.Napríklad, ak je kódujúca oblasť amplifikovaná, 3' koniec priméru by nemal byť ukončený v tretej polohe kodónu, pretože tretia poloha kodónu je náchylná k degenerácii, čo ovplyvní špecifickosť a účinnosť amplifikácie.Pri použití anexovacích primérov si pozrite tabuľku použitia kodónov, venujte pozornosť biologickej preferencii, nepoužívajte anexačné priméry na 3′ konci a použite vyššiu koncentráciu primérov (1 uM – 3 uM).
7. Sekundárna štruktúra primerov
Samotné priméry by nemali mať komplementárne sekvencie, inak sa samotné priméry zložia do vlásenkových štruktúr a táto sekundárna štruktúra ovplyvní väzbu primérov a templátov v dôsledku stérickej zábrany.Ak sa použije umelý úsudok, kontinuálne komplementárne bázy samotných primérov by nemali byť väčšie ako 3 bp.Medzi týmito dvoma primérmi by nemala existovať žiadna komplementarita, najmä by sa malo zabrániť komplementárnemu prekrývaniu 3' konca, aby sa zabránilo tvorbe primérových dimérov.Vo všeobecnosti by medzi párom primérov nemalo byť viac ako 4 po sebe idúce bázové homológie alebo komplementarita.
8. Pridajte značky alebo lokusy
5' koniec má malý vplyv na amplifikačnú špecifickosť, a preto môže byť modifikovaný bez ovplyvnenia amplifikačnej špecifickosti.Modifikácia 5' konca primeru zahŕňala: pridanie enzýmového reštrikčného miesta;Značený biotín, fluorescencia, digoxín, Eu3+ atď. Zavedenie sekvencií DNA viažucich proteín;Zavedenie mutačných miest, vloženie a chýbajúce mutačné sekvencie a vloženie promótorových sekvencií atď. Ďalšie bázy viac-menej ovplyvnia účinnosť amplifikácie a zvýšia možnosť tvorby primérového diméru, ale v ďalšom kroku je potrebné urobiť určité ústupky.Ďalšie sekvencie, ktoré neexistujú na cieľovej sekvencii, ako sú reštrikčné miesta a promótorové sekvencie, môžu byť pridané na 5' koniec priméru bez ovplyvnenia špecificity.Tieto sekvencie nie sú zahrnuté vo výpočte hodnôt Tm primérov, ale mali by byť testované na komplementaritu a vnútornú sekundárnu štruktúru.
9. Subklony
Väčšinu času je PCR len predbežné klonovanie a potom potrebujeme subklonovať cieľový fragment do rôznych vektorov, takže musíme navrhnúť ďalšie bázy pre ďalšiu operáciu v kroku PCR.
Niektoré sekvencie navrhnuté na subklonovanie sú zhrnuté nižšie.
Pridalo sa reštrikčné miesto pre reštrikčnú endonukleázu
Pridanie enzýmových restrikčných miest je najbežnejšie používanou metódou na subklonovanie produktov PCR.Vo všeobecnosti je miesto štiepenia šesť báz, okrem 5' konca miesta štiepenia je potrebné pridať 2 až 3 ochranné bázy.Počet ochranných báz, ktoré vyžadujú rôzne enzýmy, je však rôzny.Napríklad SalⅠ nevyžaduje žiadnu ochrannú základňu, EcoRⅤ vyžaduje 1 ochrannú základňu, NotⅠ vyžaduje 2 ochranné základne a Hind Ⅲ vyžaduje 3 ochranné základne.
LIC pridáva chvost
Úplný názov LIC je klonovanie nezávislé od ligácie, metóda klonovania vynájdená spoločnosťou Navogen špecificky pre jej časť vektora pET.Nosič pET pripravený metódou LIC má nekomplementárne 12-15 bázové jednovláknové lepkavé konce, ktoré dopĺňajú zodpovedajúce lepkavé konce na cieľovom inzertnom fragmente.Na účely amplifikácie by sekvencia primeru 5' vloženého fragmentu mala dopĺňať vektor LIC.3′→5′ extranektná aktivita T4 DNA polymerázy môže po krátkom čase vytvoriť jednovláknový lepkavý koniec na vloženom fragmente.Pretože produkt môže byť vytvorený len vzájomným žíhaním pripraveného inzertného fragmentu a vektora, je tento spôsob veľmi rýchly a účinný a ide o priame klonovanie.
Riadený klon TA pridať chvost
TA klonovanie nebolo schopné zacieliť fragment do vektora, takže neskôr Invitrogen zaviedol vektor, ktorý mohol zacieliť klonovanie, ktorý obsahoval štyri prominentné bázy GTGGS na jednom konci.Preto by sa pri navrhovaní PCR primérov mali zodpovedajúcim spôsobom pridať komplementárne sekvencie, aby sa fragmenty mohli „orientovať“.
Ak máte málo času, môžete vyskúšať priamu syntézu, kombinovanie génu s vektorom, čo u musekularistov nazývame syntéza génu ET.
D. In-fúzna klonovacia metóda
Nie je potrebná ligáza, nie je potrebná žiadna dlhá reakcia.Pokiaľ sa do návrhu primerov zavedie sekvencia na oboch koncoch linearizovaného vektora, potom sa produkt PCR a linearizovaný vektor pridajú do infúzneho enzýmového roztoku obsahujúceho BSA a umiestnia sa na pol hodiny pri izbovej teplote, môže sa uskutočniť transformácia.Táto metóda je obzvlášť vhodná pre konverziu veľkého objemu.
10. Zlúčiť základný náter
Niekedy sú o návrhu primérov známe len obmedzené informácie o sekvencii.Napríklad, ak je známa len aminokyselinová sekvencia, môže byť navrhnutý zlučovací primér.Zlučovací primér je zmesou rôznych sekvencií reprezentujúcich všetky rôzne možnosti báz, ktoré kódujú jednu aminokyselinu.Ak chcete zvýšiť špecifickosť, môžete sa obrátiť na tabuľku použitia kodónov na zníženie anexie podľa preferencií základného použitia rôznych organizmov.Hypoxantín môže byť spárovaný so všetkými bázami, aby sa znížila teplota žíhania priméru.Nepoužívajte pripojené bázy na 3′ konci priméru, pretože spojenie posledných 3 báz na 3′ konci je dostatočné na spustenie PCR na nesprávnom mieste.Používajú sa vyššie koncentrácie primérov (1 uM až 3 uM), pretože priméry v mnohých anexových zmesiach nie sú špecifické pre cieľový templát.
Suroviny PCRovládanie
1. Množstvo základného náteru
Koncentrácia každého priméru je 0,1 ~ 1 umol alebo 10 ~ 100 pmol.Je lepšie dosiahnuť požadovaný výsledok s najnižším množstvom základného náteru.Vysoká koncentrácia priméru spôsobí nesúlad a nešpecifickú amplifikáciu a zvýši možnosť tvorby dimérov medzi primérmi.
2. Koncentrácia primeru
Koncentrácia primérov ovplyvňuje špecifickosť.Optimálna koncentrácia primeru je všeobecne medzi 0,1 a 0,5 uM.Vyššie koncentrácie primérov vedú k amplifikácii nešpecifických produktov.
3. Teplota žíhania základného náteru
Ďalším dôležitým parametrom primérov je teplota topenia (Tm).Toto je teplota, pri ktorej je 50 % primérov a komplementárnych sekvencií reprezentovaných ako molekuly dvojvláknovej DNA.Tm je potrebná na nastavenie teploty žíhania PCR.V ideálnom prípade je teplota anelácie dostatočne nízka na to, aby sa zabezpečilo účinné spojenie primerov s cieľovou sekvenciou, ale dostatočne vysoká na zníženie nešpecifickej väzby.Primeraná teplota žíhania od 55 ℃ do 70 ℃.Teplota žíhania je všeobecne nastavená o 5 °C nižšia ako Tm základného náteru.
Existuje niekoľko vzorcov na nastavenie Tm, ktoré sa značne líšia v závislosti od použitého vzorca a sekvencie primérov.Pretože väčšina vzorcov poskytuje odhadovanú hodnotu Tm, všetky teploty žíhania sú len východiskovým bodom.Špecifickosť sa môže zlepšiť analýzou niekoľkých reakcií, ktoré postupne zvyšujú teplotu žíhania.Začnite pod odhadovanou Tm-5 °C a postupne zvyšujte teplotu žíhania v prírastku 2 °C.Vyššia teplota žíhania zníži tvorbu primérových dimérov a nešpecifických produktov.Na dosiahnutie najlepších výsledkov by dva priméry mali mať približné hodnoty Tm.Ak je rozdiel Tm párov primérov väčší ako 5 °C, priméry budú vykazovať výrazne nesprávny štart pri použití nižšej teploty žíhania v cykle.Ak sa dva priméry Tm líšia, nastavte teplotu žíhania o 5 °C nižšiu ako je najnižšia Tm.Alternatívne, na zvýšenie špecificity, možno najskôr uskutočniť päť cyklov pri teplotách žíhania navrhnutých pre vyššiu Tm, po ktorých nasledujú zvyšné cykly pri teplotách žíhania navrhnutých pre nižšiu Tm.To umožňuje získať čiastočnú kópiu cieľovej šablóny v stiesnených podmienkach.
4. Čistota a stabilita základného náteru
Štandardná čistota vlastných primerov je dostatočná pre väčšinu aplikácií PCR.Odstránenie benzoylových a izobutylylových skupín odsolením je minimálne, a preto neinterferuje s PCR.Niektoré aplikácie vyžadujú čistenie, aby sa odstránili akékoľvek sekvencie, ktoré nemajú úplnú dĺžku v procese syntézy.Tieto skrátené sekvencie sa vyskytujú, pretože účinnosť chémie syntézy DNA nie je 100%.Ide o kruhový proces, ktorý využíva opakované chemické reakcie, keď sa každá báza pridáva na vytvorenie DNA od 3′ do 5′.Môžete zlyhať v každom cykle.Dlhšie priméry, najmä tie, ktoré sú väčšie ako 50 báz, majú veľký podiel skrátených sekvencií a môžu vyžadovať purifikáciu.
Výťažok primérov je ovplyvnený účinnosťou syntetickej chémie a spôsobu čistenia.Biofarmaceutické spoločnosti, ako sú Cytology a Shengong, všetky používajú minimálnu jednotku OD na zabezpečenie celkového výstupu oligonukleozidu.Vlastné primery sa dodávajú vo forme suchého prášku.Priméry je najlepšie znovu rozpustiť v TE tak, aby konečná koncentrácia bola 100 μM.TE je lepšia ako deionizovaná voda, pretože pH vody je často kyslé a spôsobí hydrolýzu oligonukleozidov.
Stabilita primerov závisí od podmienok skladovania.Suchý prášok a rozpustené základné nátery by sa mali skladovať pri teplote -20 °C.Priméry rozpustené v TE v koncentráciách vyšších ako 10 μM možno stabilne skladovať pri -20 °C po dobu 6 mesiacov, ale možno ich skladovať iba pri izbovej teplote (15 °C až 30 °C) menej ako 1 týždeň.Suché práškové základné nátery je možné skladovať pri -20 C minimálne 1 rok a pri izbovej teplote (15 C až 30 C) až 2 mesiace.
5. Enzýmy a ich koncentrácie
V súčasnosti používaná Taq DNA polymeráza je v podstate enzým génového inžinierstva syntetizovaný koliformnými baktériami.Množstvo enzýmu potrebného na katalýzu typickej PCR reakcie je približne 2,5 U (vzťahuje sa na celkový reakčný objem 100 ul).Ak je koncentrácia príliš vysoká, môže to viesť k nešpecifickej amplifikácii;ak je koncentrácia príliš nízka, množstvo syntetického produktu sa zníži.
6. Kvalita a koncentrácia dNTP
Kvalita dNTP úzko súvisí s koncentráciou a účinnosťou PCR amplifikácie.Prášok dNTP je zrnitý a jeho variabilita stráca svoju biologickú aktivitu, ak sa nesprávne skladuje.Roztok dNTP je kyslý a mal by sa používať vo vysokej koncentrácii, s 1M NaOH alebo 1M Tris.HCL tlmivým roztokom na úpravu jeho pH na 7,0 ~ 7,5, malé množstvo čiastkového balenia, mrazené skladovanie pri -20 °C.Viacnásobné zmrazenie-rozmrazenie degraduje dNTP.V PCR reakcii by dNTP mala byť 50 ~ 200 umol/l.Predovšetkým by sa mala venovať pozornosť tomu, aby koncentrácia štyroch DNTPS bola rovnaká (rovnaký molový prípravok).Ak je koncentrácia ktorejkoľvek z nich odlišná od ostatných (vyššia alebo nižšia), spôsobí to nesúlad.Príliš nízka koncentrácia zníži výťažok produktov PCR.dNTP sa môže kombinovať s Mg2+ a znižovať koncentráciu voľného Mg2+.
7. Templátová (cieľový gén) nukleová kyselina
Množstvo a stupeň purifikácie templátovej nukleovej kyseliny je jedným z kľúčových spojení pre úspech alebo zlyhanie PCR.Tradičné metódy purifikácie DNA zvyčajne používajú SDS a proteázu K na štiepenie a likvidáciu vzoriek.Hlavné funkcie SDS sú: rozpúšťanie lipidov a proteínov na bunkovej membráne, čím dochádza k deštrukcii bunkovej membrány rozpustením membránových proteínov a disociácia jadrových proteínov v bunke, SDS sa môže spájať aj s proteínmi a precipitovať;Proteáza K môže hydrolyzovať a tráviť proteíny, najmä históny viazané s DNA, a potom použiť organické rozpúšťadlo fenol a chloroform na extrakciu proteínov a iných bunkových zložiek a použiť etanol alebo izopropylalkohol na vyzrážanie nukleovej kyseliny.Extrahovaná nukleová kyselina môže byť použitá ako templát pre PCR reakcie.Pre všeobecné klinické detekčné vzorky možno použiť rýchlu a jednoduchú metódu na rozpustenie buniek, lyzáciu patogénov, štiepenie a odstránenie proteínov z chromozómov na uvoľnenie cieľových génov a priamo na použitie na PCR amplifikáciu.Extrakcia templátu RNA zvyčajne používa metódu guanidín izotiokyanát alebo proteázu K, aby sa zabránilo RNáze degradovať RNA.
8. Koncentrácia Mg2+
Mg2+ má významný vplyv na špecifickosť a výťažok PCR amplifikácie.Vo všeobecnej PCR reakcii, keď je koncentrácia rôznych dNTP 200 umol/l, vhodná koncentrácia Mg2+ je 1,5 ~ 2,0 mmol/l.Koncentrácia Mg2+ je príliš vysoká, znižuje sa špecificita reakcie, dochádza k nešpecifickej amplifikácii, príliš nízka koncentrácia zníži aktivitu Taq DNA polymerázy, čo vedie k redukcii produktov reakcie.
Ióny horčíka ovplyvňujú niekoľko aspektov PCR, ako je aktivita DNA polymerázy, ktorá ovplyvňuje výťažok;Ďalším príkladom je žíhanie primérov, ktoré ovplyvňuje špecifickosť.dNTP a templát sa viažu na ión horčíka, čím sa znižuje množstvo voľného iónu horčíka potrebného na aktivitu enzýmu.Optimálna koncentrácia horčíkových iónov sa líši pre rôzne páry primérov a templátov, ale typická východisková koncentrácia pre PCR s 200 μM dNTP je 1,5 mM (poznámka: Pre kvantitatívnu PCR v reálnom čase použite 3 až 5 mM roztok horčíkových iónov s fluorescenčnou sondou).Vyššie koncentrácie voľných iónov horčíka zvyšujú výťažok, ale tiež zvyšujú nešpecifickú amplifikáciu a znižujú vernosť.Na stanovenie optimálnej koncentrácie sa titrácie iónov horčíka uskutočňovali v prírastkoch 0,5 mM od 1 mM do 3 mM.Na zníženie závislosti od optimalizácie horčíkových iónov možno použiť DNA polymerázu Platinum Taq.Platinová Taq DNA polymeráza je schopná zachovať funkciu v širšom rozsahu koncentrácií horčíkových iónov ako Taq DNA polymeráza, a preto vyžaduje menšiu optimalizáciu.
9. Prísady podporujúce Pcr
Optimalizácia teploty žíhania, dizajnu primeru a koncentrácie horčíkových iónov postačuje na vysoko špecifickú amplifikáciu väčšiny templátov;niektoré šablóny, vrátane šablón s vysokým obsahom GC, si však vyžadujú dodatočné opatrenia.Aditíva, ktoré ovplyvňujú teplotu topenia DNA, poskytujú ďalší spôsob, ako zlepšiť špecifickosť produktu a výťažok.Na dosiahnutie najlepších výsledkov je potrebná úplná denaturácia šablóny.
Okrem toho sekundárna štruktúra zabraňuje väzbe priméru a predlžovaniu enzýmu.
PCR aditíva, vrátane formamidu, DMSO, glycerínu, betaínu a PCRx Enhancer Solution, zvyšujú amplifikáciu.Ich možným mechanizmom je zníženie teploty topenia, čím napomáhajú žíhaniu primérov a napomáhajú predlžovaniu DNA polymerázy cez oblasť sekundárnej štruktúry.Roztok PCRx má ďalšie výhody.Pri použití s Platinum Taq DNA polymerázou a Platinum Pfx DNA polymerázou je potrebná minimálna optimalizácia horčíkových iónov.Platinová technika je teda kombinovaná s aditívom na zvýšenie špecifickosti a zároveň zníženie závislosti od tretieho prístupu, optimalizácie horčíkových iónov.Na dosiahnutie najlepších výsledkov by sa mala optimalizovať koncentrácia aditív, najmä DMSO, formamid a glycerol, ktoré inhibujú Taq DNA polymerázu.

10. Horúci štart
Hot start PCR je jednou z najdôležitejších metód na zlepšenie špecifickosti PCR okrem dobrého návrhu primeru.Hoci optimálna teplota predĺženia Taq DNA polymerázy je 72 °C, polymeráza zostáva aktívna pri izbovej teplote.Nešpecifické produkty sú teda produkované, keď je udržiavacia teplota nižšia ako teplota žíhania počas prípravy PCR reakcie a na začiatku tepelného cyklu.Po vytvorení sú tieto nešpecifické produkty efektívne zosilnené.Hot-start PCR je obzvlášť účinná, keď sú miesta použité na návrh primérov obmedzené umiestnením genetických prvkov, ako sú miestne cielené mutácie, expresné klonovanie alebo konštrukcia a manipulácia genetických prvkov používaných na DNA inžinierstvo.
Bežnou metódou na obmedzenie aktivity Taq DNA polymerázy je pripraviť reakčný roztok PCR na ľade a umiestniť ho do predhriateho PCR prístroja.Táto metóda je jednoduchá a lacná, ale nedokončuje aktivitu enzýmu, a preto úplne neodstraňuje amplifikáciu nešpecifických produktov.
Tepelná aktivácia oneskoruje syntézu DNA inhibíciou základnej zložky, kým prístroj PCR nedosiahne denaturačnú teplotu.Väčšina metód manuálnej tepelnej iniciácie, vrátane oneskoreného pridania Taq DNA polymerázy, je ťažkopádna, najmä pre aplikácie s vysokou priepustnosťou.Iné metódy tepelného nanášania používajú voskový štít na uzavretie základnej zložky, vrátane horčíkových iónov alebo enzýmov, alebo na fyzikálnu izoláciu reaktívnych zložiek, ako sú templáty a pufre.Počas tepelného cyklu sa rôzne zložky uvoľňujú a miešajú sa, keď sa vosk topí.Podobne ako manuálna metóda horúceho štartu je metóda voskového štítu ťažkopádna a náchylná na kontamináciu a nie je vhodná pre aplikácie s vysokou priepustnosťou.
Platinová DNA polymeráza je vhodná a efektívna pre automatický hot start PCR.Platinová Taq DNA polymeráza pozostáva z rekombinantnej Taq DNA polymerázy kombinovanej s monoklonálnou protilátkou proti Taq DNA polymeráze.Protilátky sú formulované pomocou PCR, aby inhibovali aktivitu enzýmu počas dlhšieho udržiavania teploty.Taq DNA polymeráza sa uvoľnila do reakcie počas 94 °C izolácie v kroku denaturácie, čím sa obnovila plná polymerázová aktivita.Na rozdiel od chemicky modifikovanej Taq DNA polymerázy na tepelnú iniciáciu, enzým Platinum nevyžaduje predĺženú izoláciu pri 94 °C (10 až 15 minút) na aktiváciu polymerázy.S PlatinumTaq DNA polymerázou sa 90 % aktivity Taq DNA polymerázy obnovilo po 2 minútach pri 94 °C.

11. Nest-PCR
Postupné cykly amplifikácie s použitím vnorených primérov môžu zlepšiť špecificitu a citlivosť.Prvé kolo je štandardné zosilnenie 15 až 20 cyklov.Malá frakcia počiatočného amplifikačného produktu sa zriedila 100 až 1000-krát a pridala sa do druhého kola amplifikácie na 15 až 20 cyklov.Alternatívne môže byť veľkosť počiatočného amplifikovaného produktu určená gélovou purifikáciou.V druhom kole amplifikácie sa používa vnorený primér, ktorý sa môže viazať na cieľovú sekvenciu vo vnútri prvého priméru.Použitie nested PCR znižuje možnosť amplifikácie viacerých cieľových miest, pretože existuje málo cieľových sekvencií komplementárnych k obom súborom primérov.Rovnaký celkový počet cyklov (30 až 40) s rovnakými primérmi amplifikoval nešpecifické miesta.Nested PCR zvyšuje citlivosť obmedzených cieľových sekvencií (napr. zriedkavé mrnas) a zlepšuje špecifickosť náročných PCRS (napr. 5′ RACE).
12. Zostupná PCR
Zostupná PCR zlepšuje špecificitu použitím tesných podmienok nasadenia počas prvých niekoľkých cyklov PCR.Cyklus začína pri teplote žíhania približne o 5 °C vyššej ako je odhadovaná Tm, potom sa každý cyklus zníži o 1 °C až 2 °C, kým teplota žíhania neklesne pod Tm 5 °C.Amplifikovaný bude iba cieľový templát s najvyššou homológiou.Tieto produkty pokračujú v expanzii v nasledujúcich cykloch a vytláčajú zosilnené nešpecifické produkty.Zostupná PCR je užitočná pre metódy, kde stupeň homológie medzi primérom a cieľovým templátom nie je známy, ako je napríklad AFLP DNA fingerprinting.
Súvisiace súpravy PCR
Hrdina 2× PCRTMMix systém má vyššiu toleranciu voči inhibítorom PCR ako bežný systém PCR Mix a ľahko si poradí s PCR amplifikáciou rôznych komplexných templátov.Vďaka jedinečnému reakčnému systému a vysoko účinnému Taq Hero má PCR reakcia vyššiu účinnosť amplifikácie, špecifickosť a citlivosť.
Vyššia účinnosť zosilnenia
Má 5'→3' DNA polymerázovú aktivitu a 5'→3' exonukleázovú aktivitu, bez 3'→5' exonukleázovej aktivity.
Real Time PCR Easyᵀᴹ-SYBR Green I Kit
Špecifický – optimalizovaný pufor a enzým Taq s horúcim štartom môžu zabrániť nešpecifickej amplifikácii a tvorbe primérového diméru
Vysoká citlivosť—môže zistiť nízke kópie šablóny
Súprava využíva unikátne reverzné transkripčné činidlo Foregene a Foregene HotStar Taq DNA polymerázu v kombinácii s jedinečným reakčným systémom na efektívne zlepšenie účinnosti amplifikácie a špecifickosti reakcie.
Čas odoslania: máj-09-2023



